
Innate sensing of vita-PAMPs, signatures of microbial viability
What are vita-PAMPs?
We discovered that the innate immune system can detect and respond to microbial viability, the quintessential basis of infectivity. Vita-PAMPs is a term we coined to refer to microbial structures that signify microbial viability. Pathogen-associated molecular patterns (PAMPs) are present in microbes and absent from host tissues. They alert the innate immune system. Examples of PAMPs include lipopolysaccharide (LPS), a major cell wall component of Gram-negative bacteria, lipoteichoic acid present in Gram-positive bacterial cell walls, or DNA from viruses. On the other hand, bacterial messenger RNA (mRNA) and c-di-adenosine mono-phosphate (c-di-AMP) are vita-PAMPs uniquely associated with live microorganisms.
A question that we get asked is why rely on vita-PAMPs when live pathogens announce their presence by expressing virulence factors that facilitate invasion and damage their hosts. Virulence factor expression is transient serving a specific purpose such as breaching an epithelial barrier. Once microbes gain access to sterile tissues, relying on vita-PAMPs is a fail-safe strategy for eliminating the threat. The ability to discriminate between live and dead microbes also tailors the immune response to the threat level it encounters. It avoids collateral damage to the host from an overly exuberant response (See Sander L.E. et al. Nature 2011; Moretti, J. et al. Cell 2017; Blander and Sander, Nat Rev Immunol 2012).
A distinct innate immune response to vita-PAMPs
When Gram-negative bacteria invade the cytosol of cells, lipopolysaccharide (LPS) from their cell walls directly binds to and activates the cytosolic LPS sensors murine caspase-11 and human caspase-4. This results in noncanonical activation of the NLRP3 inflammasome leading to death of the infected cell and secretion of IL1-beta to mobilize immune responses. Non-virulent bacteria do not invade cells, yet their presence in tissue is a threat that warrants immune defense mobilization. Accordingly, we found that various avirulent Gram-negative bacteria can also trigger inflammasome activation as long as they are viable. We found that the NOD, LRR and pyrin domain containing 3 (NLRP3) inflammasome is activated in response to live Gram-negative bacteria upon detection of the vita-PAMP bacterial mRNA (See Sander L.E. et al. Nature 2011; Blander and Sander, Nat Rev Immunol 2012; Moretti J. et al. Nat Immunol 2022).
The innate immune response also has other ways of mounting a specific response only to live bacteria. A second vita-PAMP we identified is the second messenger cyclic (c)-diAMP from live Gram-positive bacteria. c-diAMP engages an intracellular protein called STING on the endoplasmic reticulum (ER). We found that STING engagement by c-di-AMP triggers a form of ER stress in responding phagocytes, which then mobilize autophagy to resolve the ER stress. This autophagy of the ER is called ER-phagy and it serves two functions. First, it enables the phagocytes to survive and do their job, and second, it delivers STING from ER membranes to autophagosomes where it can signal type-I interferon production (See Moretti, J. et al. Cell 2017; Moretti and Blander, Autophagy 2018). None of these elaborate phagocyte responses are mobilized to killed bacteria.

PAMP = dead microbe
PAMP + vita-PAMP = live/viable microbe
Two vita-PAMPs: Bacterial messenger RNA (mRNA), essential for the viability of all bacteria, and the cyclic dinucleotide adenosine mono-phosphate (c-di-AMP) made by growing Gram-positive bacteria.

Electron micrographs showing pyroptosis in mouse macrophages at 12 hours only after stimulation with live avirulent Escherichia coli. This pyroptosis is mediated by non-canonical NLRP3 inflammasome activation and is dependent on the detection and response to bacterial RNA and LPS (see Sander LE et al. Nature 2011; Moretti J et al. Cell 2017).

Model of non-canonical NLRP3 inflammasome activation upon coincident detection of the vita-PAMP bacterial mRNA and the classic PAMP LPS. Based on Sander LE et al. Nature 2011; Moretti J. et al. Nat Immunol 2021. Work by others is also highlighted. Created with BioRender.com
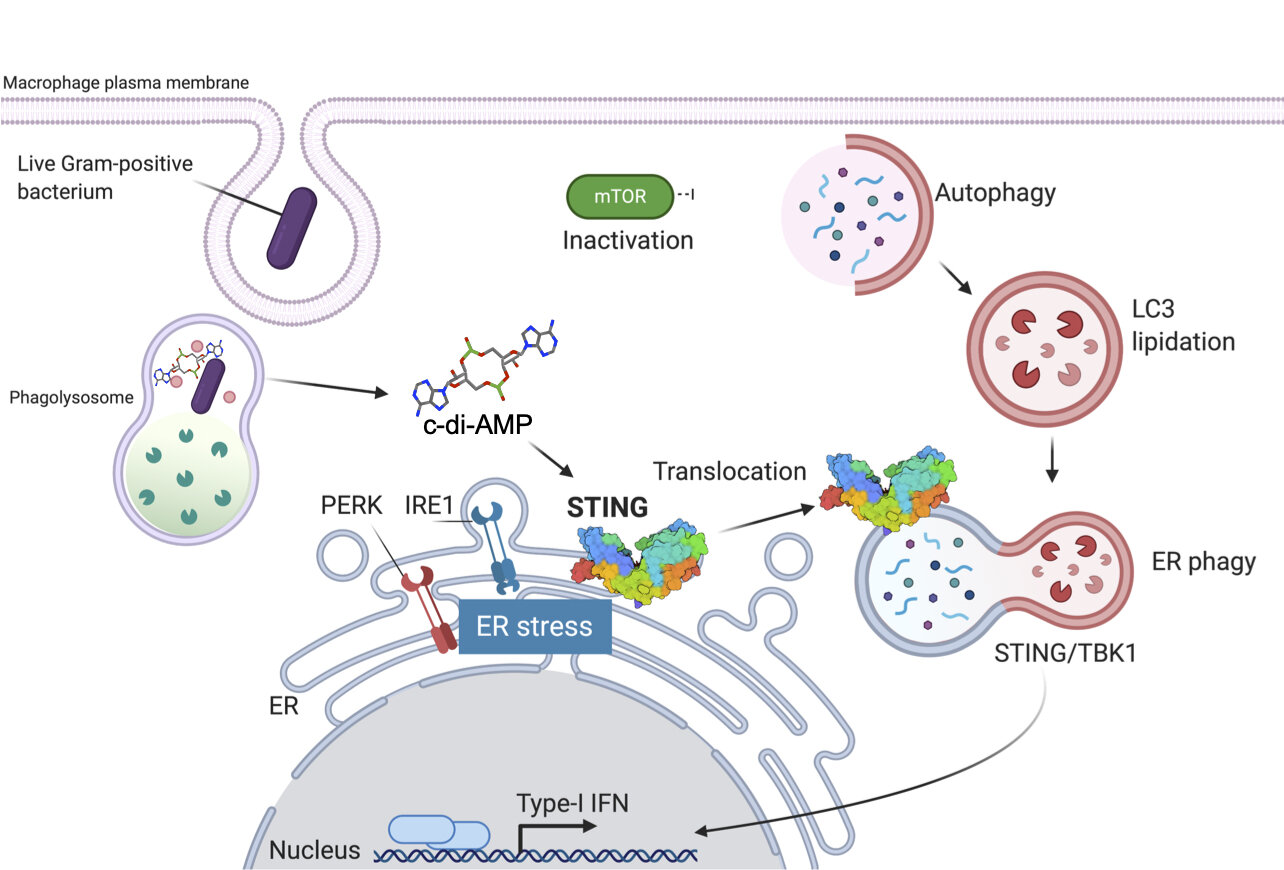
Schematic of the macrophage innate response to the vita-PAMP c-di-AMP from Gram-positive bacteria. See Moretti J et al. Cell 2017. Created with BioRender.com
Exploiting vita-PAMPs in vaccines

The vita-PAMP bacterial mRNA drives augmented follicular T helper cell and antibody responses. Detection of the vita-PAMP bacterial mRNA by patrolling monocytes in the context of Gram-negative bacteria triggers TRIF signaling and IRF3-dependent interferon (IFN) beta production, which engages IFNAR to license activation of the inflammasome and lead to the production of the alert cytokine interleukin-1 (IL-1) beta. IFN-beta and IL-1 beta engage their respective receptors on CD4 T cells instructing their differentiation into follicular T helper cells (TFH), known for their roles in helping B cells make antibodies. This orchestrated series of events from detection of a vita-PAMP to the adaptive humoral immune response illustrates how key first events in innate immunity shape the quality of the adaptive immune response. See Barbet G et al. Immunity 2018.
Live attenuated vaccines have proven to be the most efficient human vaccines. When compared to their dead counterparts, live vaccines elicit superior protection and lasting immune memory. But despite the efficacy of live vaccines, concerns over their safety have limited their use especially in the very young, the elderly and immunocompromised. Preservation and delivery of live vaccines, especially to impoverished areas in developing countries, is difficult and expensive. Understanding the molecular basis for the efficacy of live vaccines is important because it would solve these limitations leading to the development of vaccines that are safe and afford the same protection as live vaccines.
Using experimental models of vaccination in mice, we delineated distinct connections from the innate pathways engaged by vita-PAMPs to the adaptive humoral response. Supplementing a dead vaccine with bacterial RNA (what we call a vita-vaccine) augments its performance through the innate pathways engaged specifically by live bacteria (See Sander L.E. et al. Nature 2011). A vita-vaccine performed similarly to a live vaccine in eliciting a follicular T helper cell response (that helps B cells), germinal center formation, and B cell isotype class switching (See Barbet G. et al. 2018 Immunity). Our work indicates that vita-PAMPs underlie the superior performance of live vaccines. Defining the molecular and cellular context in which vita-PAMPs are recognized will further inform the design of safe and efficacious vaccines (See Barbet and Blander Curr Opin Pharmacol 2018).